|
京都大学生態学研究センター、熊本大学大学院先端科学研究部附属生物環境農学国際研究センター、千葉大学大学院薬学研究院、名城大学農学部、森林総合研究所、理化学研究所環境資源科学研究センター、琉球大学熱帯生物圏研究センター、静岡大学農学部からなる研究チームは、多年生植物のオオバコの種子がダンゴムシの糞に含まれる化学物質を感知して発芽を一時的に止め、ダンゴムシによる食害を回避する仕組みを発見しました(図1)。ダンゴムシの糞中に含まれる「トレハロース」と「アブシジン酸(ABA)」が発芽を一時的に抑制すること、そしてそれらの成分が水で洗い流されると発芽が再開することが明らかになりました。さらに野外調査では、ダンゴムシの糞が存在する環境では、雨天時にオオバコ種子の発芽が集中し、ダンゴムシによる食害が起こりにくいことも確認されました。これらの結果は、オオバコの種子がダンゴムシの活動が活発な晴天時には発芽を抑え、活動が低下する雨天時に発芽を促すことで、食害を免れやすくなることを示唆しています。植物の種子はこれまで、光や温度などの環境刺激に応じて発芽時期を調整することが知られていましたが、本研究は、種子が植食者由来の刺激にも反応し、食害を回避できることを初めて明らかにしました。 図 1.研究の概要図.ダンゴムシの糞に含まれる2つの化学物質がダンゴムシの活動が低下する安全な雨天時の種子の発芽を促進する. |

1.背景
発芽した直後の実生は、防御機構がまだ十分に発達しておらず、草食動物に食べられやすい発生段階にあります。特に小さな芽生えでは、わずかな食害でも致命的なダメージとなることがあります。これまで、植物の種子が光や温度といった物理環境や競争者となる他の植物の存在に応じて発芽を調整することは知られていましたが、「動物の存在に関わる刺激」に反応できるかどうかは明らかにされていませんでした。
研究チームは、多年草のオオバコ(Plantago asiatica)と、その芽生えを高頻度で食べるダンゴムシ(Armadillidium vulgare)およびワラジムシ(Porcellio scaber)に注目しました。オオバコの群落内では、ダンゴムシが親株の古い葉を頻繁に食べるだけでなく、種子から出たばかりの幼根を食べて芽生えを枯死させます。また、事前の観察から親株の葉や芽生えを食べたダンゴムシは、周囲に多くの糞を蓄積することも分かりました(図2)。

図 2. (a)ダンゴムシに食べられた葉(白矢印).(b)蓄積されたダンゴムシの糞(黄矢印)と散布された種子(黄色矢印).
これらの観察から、研究チームは「オオバコの種子が、ダンゴムシの糞を感知し、発芽を一時的に抑制するといった、芽生え時の食害を回避する発芽応答が自然選択により進化しているのではないか」と予測し、ダンゴムシ類の糞が種子の発芽や芽生えの生存に与える影響について調査しました。
2.研究手法・成果
私たちはまず、ダンゴムシの糞の存在がオオバコの発芽に影響するかどうかを調べました。シャーレにオオバコの種子を単独で播いた場合と、ダンゴムシやワラジムシの糞と一緒に播いた場合とで、発芽率や発芽速度に違いがあるかを比較しました。1日間絶食させたダンゴムシに古くなったオオバコの葉を与え、排出された糞を集めて実験に用いました。その結果、オオバコの種子を単独で播いた条件(コントロール)では、約97%と高い発芽率を示したのに対して、ダンゴムシの糞と一緒に播く条件では、約20%と発芽率は大きく低下しました(図3a)。また、一部の発芽した種子も発芽までにかかる日数が約8日と、コントロールにおける4-5日に比べて、遅くなることが分かりました。
図3.(a)ワラジムシとダンゴムシの糞、および磨り潰したオオバコの葉が種子の発芽率に与える影響と、(b)それらを水で洗浄した際の発芽率.アルファベットの違いと**は、統計的に有意な差異があることを示す.
さらに、降雨によりダンゴムシの糞や化学成分が流される状況を模倣し、水洗によって種子周辺から糞を除去したところ、発芽していなかった種子が約2日後に正常に発芽することが明らかになりました(図3b)。これらの実験の結果から、オオバコの種子は近くにダンゴムシ等の糞がある場合には、糞が水で除去されるまで発芽を抑制することが分かりました。
研究チームは、このようなダンゴムシの糞に応じた発芽タイミングの変化が、実生の食害回避に寄与しているかどうかを検証するため、糞の有無を操作した野外実験を行いました。オオバコの種子400粒を「ダンゴムシの糞がある条件」と「糞がない条件」にそれぞれ設置し(合計800粒)、連続撮影システム(タイムラプスカメラ)を用いて、発芽や食害の経過を詳細に20日間記録しました。得られた画像を天候データと照合したところ、糞のない条件では発芽は晴天・雨天でほぼ同程度に起こったのに対し、糞のある条件では発芽が雨天時に集中することが明らかになりました。これは、室内実験で得られた「糞を水で洗うと発芽が再開する」という結果と一致しており、糞に含まれる発芽抑制物質が雨によって洗い流されることで発芽が促進されることを示唆しています。
加えて、降雨時には地表を徘徊するダンゴムシの密度が著しく低下することも確認されました。これは、ダンゴムシが雨水による溺死を避けるため、石や草の上などの高所へ避難する行動によるものと考えられます。その結果、降雨時にはダンゴムシによる芽生えの食害が減少し、糞を設置した条件では発芽後の幼根が食べられる割合が低下しました。その結果、糞のある条件での実生の最終的な生存率は、糞のない条件の数倍に達しました(図4)。別の年に行った実験でも同様に、ダンゴムシの糞の存在が実生の生存率を高めることが確認されました。特に、糞の効果はダンゴムシの生息密度が低い場所よりも、高密度に生息する場所でより顕著に現れることも分かりました。
これらの結果から、オオバコの種子は、ダンゴムシの糞を化学的に感知して発芽タイミングをダンゴムシの活動量の低い雨天時にまで遅らせていること、この発芽タイミングの変化によりダンゴムシによる食害を免れていることが分かりました。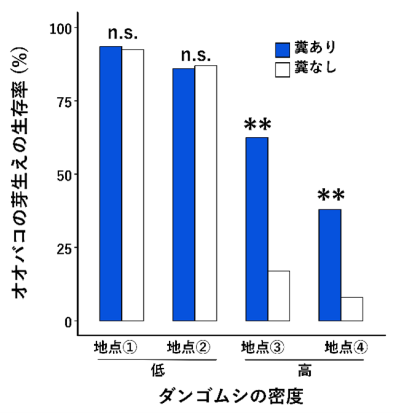
図 4.ダンゴムシの生息密度が異なる地域における、ダンゴムシの糞がオオバコの芽生えの生存率に与える影響.n.s. は統計的に有意な差異がないことを、**は統計的に有意な差異があることを示す.
最後に、研究チームは詳細な化学分析を実施し、この現象にどのような化学物質が関与しているのかを特定しました。その結果、アブシジン酸(ABA)という植物ホルモンの一種がダンゴムシの糞内から検出され、発芽抑制の活性成分として特定されました。ABAの働きを弱める試薬PANMe(フェニル酢酸メチルエステル)とフルリドン(Fluridone)という2種類の阻害剤を用いることで、ABAの作用機構を調べました。PANMe は、ABAが種子の中で働くのを妨げる「作用阻害剤」として機能します。つまり、すでに存在するABAの効果を打ち消す物質です。一方、フルリドンは、ABAが植物体内で合成されるのを防ぐ「生合成阻害剤」で、ABAそのものの蓄積を抑える働きをします。実験の結果、PANMeとフルリドンは、共に糞による発芽抑制効果を解除することが分かりました。このことから、糞内のABAの受容と種子によるABA合成の両方が発芽抑制に関与していることが明らかになりました。
しかし、実際に糞内や種子内のABA濃度は非常に微量であり、糞中と同程度の濃度のABAをオオバコの種子に添加しても、発芽が抑制されないことが分かりました。これにより、種子の発芽抑制にはABA単独の働きでは不十分であることが示されました。
そこで、さらに詳細な分析を進めたところ、もう一つの発芽抑制成分として多糖類の一種であるトレハロースが特定されました。トレハロースは、植物体内でABAの活性を相乗的に高めることが知られています。ABA濃度とトレハロース濃度を複数段階に操作した溶液を用いてオオバコの種子に添加したところ、糞内と同様の低いABA濃度であっても、トレハロースと併せて添加すると顕著に発芽が抑制されることが分かりました。さらに、トレハロースの発芽抑制効果は、構造異性体であるL-トレハロース(自然界には存在しない形態)では観察されませんでした。これらの結果から、種子の発芽抑制はトレハロースによる浸透圧変化などの物理的作用によるものではなく、種子がトレハロースそのものをシグナル分子として受容していること、すなわち、種子にトレハロース分子の受容体が存在する可能性が示唆されました。
ABAが植物ホルモンの一種であること、また先行研究でダンゴムシ類がトレハロースの消化・吸収を苦手とすることが知られていることから、ABAとトレハロースは親植物の葉由来であり、ダンゴムシの消化管を経て糞中に蓄積したものであると推測されました。実際、ダンゴムシの糞の代わりに人為的にすりつぶしたオオバコの葉を種子に添加する実験でも、糞を用いた場合と同様に種子の発芽遅延と発芽再開が観察されました(図3)。これらの結果から、親植物の化学物質が動物を介して子の発芽に影響を与える、いわば動物を媒介とした「植物の世代間情報伝達」の可能性が示唆されました。
3.波及効果、今後の予定
本研究により、植物の種子が動物の排泄物に含まれる化学物質を情報源として利用するという、新たな生態的相互作用の存在が明らかになりました。自然界では多種多様な動物の糞が豊富に存在していることから、糞中の化学物質を利用した発芽制御の仕組みは、オオバコに限らず、他の多くの植物にも見られる可能性があります。
一方で、今回対象としたダンゴムシ類(Armadillidium vulgare, Porcellio scaber)は外来種であることから、今回明らかになったオオバコとの関係性がどのように形成されたのかという新たな疑問も生じます。日本には在来のダンゴムシも生息しており、もともとはオオバコが在来種ダンゴムシに対して進化させた防御機構が、結果的に外来種に対しても有効に働いた可能性が考えられます。今回発見された植食者の糞に対する発芽応答が、真に自然選択の結果として生じたのかどうかは未解明のままです。今後は、在来種と外来種のダンゴムシ間での相互作用の違いや、この現象が他の植物種にも普遍的に見られるかどうかを検証することで、種子と動物の新たな関係性が見えてくるものと考えられます。
4.研究プロジェクト
本研究は、日本学術振興会科学研究費補助金(18H05487, 20H00422, 18K19353, 19H03295)の支援を受けて実施されました。
<研究者のコメント>
植物が種子の段階から動物の糞に由来する化学的刺激に反応し、その反応が生存率を高める機能を持つこと、さらに糞が取り除かれるとすぐに発芽するという種子の柔軟な環境応答にはとても驚かされました。今後は、このような反応がどのように進化してきたのか、あるいは他の植物種でも一般的に見られる現象なのかを調査し、種と糞の相互作用の全容に迫りたいと考えています。(山尾 僚)
最初に山尾さんからこの現象の話を聞いたとき、正直なところ「そんなことが本当にあるのか」と半信半疑でした。植物が動物の糞に含まれる化学物質を手がかりに発芽を調整するという現象は、これまでの常識では想像しにくかったからです。しかし研究が進むにつれて、この現象がこれまで誰も気づいてこなかった生態学的な大発見であることを実感しました。今後は、植物と動物の間の化学的コミュニケーションがどのように進化し、生態系の中でどのような役割を果たしているのかについて、より深く探究していく研究領域を育てていくことが重要だと考えています。(澤 進一郎)
トレハロースは糖の一種です。通常、糖は捕食者にとって栄養となるはずですが、オオバコはダンゴムシが消化できない糖であるトレハロースを作りだします。そして、ダンゴムシの体内を通過させることで、その糞中にトレハロースを移動させています。実際に、私が担当したダンゴムシの糞の分析では、見事にトレハロース以外の糖は観測されませんでした。このトレハロースが種子の発芽を抑制するという事実に、生命システムの巧みさと美しさを強く感じます。(石川 勇人)
<論文タイトルと著者>
タイトル:Isopod-feces-mediated shifts in germination timing enhance seedling establishment
ダンゴムシの糞が植物の発芽タイミングを変化させ、実生の定着を促す
著者:山尾僚*¹, 石川勇人*², 武川正嗣, 中嶋なな子, 大﨑晴菜*³, 向井裕美*⁴, 菅野 裕理, 瀬尾 光範*⁵, 轟 泰司, 竹内純*⁶, 澤 進一郎*⁷
掲載誌:New Phytologist DOI:10.1111/nph.70750
URL:https://nph.onlinelibrary.wiley.com/doi/10.1111/nph.70750
*1京都大学生態学研究センター教授
*2千葉大学大学院薬学研究院教授
*3名城大学農学部助教
*4森林総合研究所主任研究員
*5琉球大学熱帯生物圏研究センター教授(兼)理化学研究所環境資源科学研究センター客員主管研究員
*6静岡大学農学部教授
*7熊本大学大学院先端科学研究部附属生物環境農学国際研究センター教授
