|
琉球大学大学院理工学研究科 博士後期課程の城間 大輝 大学院生と同大学教育学部/大学院理工学研究科の富永 篤 教授,同大学大学院医学研究科の松波 雅俊 学内講師,愛知教育大学教育学部の島田 知彦 准教授,京都大学大学院地球環境学堂の西川 完途 教授,同大学大学院人間・環境学研究科の松井 正文 名誉教授らの研究チームによる研究成果が,アメリカの学術雑誌「Journal of Heredity」誌に掲載されました.本研究では,ゲノムワイド解析によりアカハライモリの複雑な集団構造と,本種に含まれる多様な遺伝グループの形成過程で,グループ間の交雑が重要な役割を担っていたことを明らかにしました. <発表のポイント> ◆新規性 ◆社会的意義/将来の展望 |

<発表概要>
琉球大学大学院理工学研究科 博士後期課程の城間 大輝 大学院生と同大学教育学部/大学院理工学研究科の富永 篤 教授,同大学大学院医学研究科の松波 雅俊 学内講師,愛知教育大学教育学部の島田 知彦 准教授,京都大学大学院地球環境学堂の西川 完途 教授および同人間・環境学研究科の松井 正文 名誉教授,龍谷大学農学部/慶應義塾大学先端生命科学研究所の永野 惇 教授からなる研究グループは,日本固有の両生類であるアカハライモリについて,次世代シーケンサーを用いた遺伝解析により詳細な遺伝的集団構造(*1)を明らかにしました.さらに,集団間の交雑(異なる種や系統間で生じる交配)や遺伝子浸透(*2)が複雑な集団構造の形成に影響を及ぼしていることを示しました.本研究の成果は,アカハライモリの分類・系統・集団構造に関する今後の研究の基盤になるとともに,生物の多様化プロセスにおける交雑の役割について理解するうえで重要な知見となります.
研究の背景
進化生物学の最近の進歩により,種間・系統間の交雑と遺伝子浸透が種の多様化において重要な役割を果たしてきたことが指摘されてきています.交雑や遺伝子浸透は新たな対立遺伝子(*3)の組み合わせや対立遺伝子頻度の変化を生み出し,様々な進化的な結果をもたらすことがあります.アカハライモリは,本州,四国,九州とその周辺離島に広く分布する日本固有の両生類です.本種の地理的変異に関する先行研究としては,形態・行動学的解析によるもの(図1)や,一部の遺伝子領域のみを調べた遺伝学的解析によるものがあり,いずれの研究でも複数の地域グループが存在していることが知られていました.それらの地域グループの地理的区分は,異なる調査手法から得られた結果にも関わらず概ね一致しているものの,一部食い違う部分もありました.

また,地域グループの分布の境界地域では,交雑個体が主に生息する地域である交雑帯(*4)が確認されており,その交雑帯を介した地域グループ間の遺伝子浸透が確認されていました.このように,多様な地理的グループを内包する本種は,生物の多様化プロセスにおける交雑の影響を評価するのに適しています.そこで,次世代シーケンサーを用いて,核に含まれるDNAの全領域を対象に遺伝解析を実施し,地域グループ間の集団構造と遺伝子浸透の実態解明を目指しました.
研究内容
本研究では,日本列島を広くカバーする28地点から得られた合計124個体の標本を対象に,従来調べられていたミトコンドリアシトクロムb領域(*5)に加えて,ddRAD-seq(*6)という手法により得られた16,273 SNPs(*7)に基づく遺伝的集団構造を調べました (図2).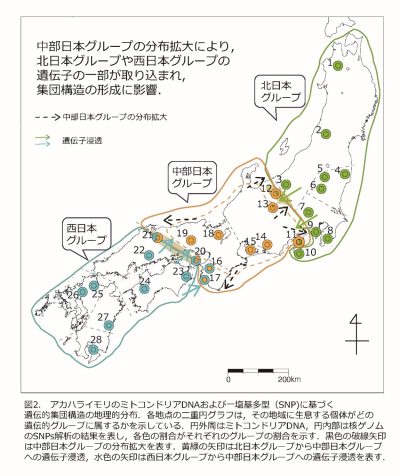
その結果,本種は,東北・関東・伊豆半島の南部に分布する北日本グループ,中部・近畿に分布する中部日本グループ,中国・四国・九州に分布する西日本グループの3つの大きなグループに分けられ,さらに各グループの中にも,これまでの先行研究で知られていた地域グループに相当する複数のサブグループが存在することが明らかになりました(図3 A B).また,グループ間の分布の境界地域では交雑集団が確認されました.
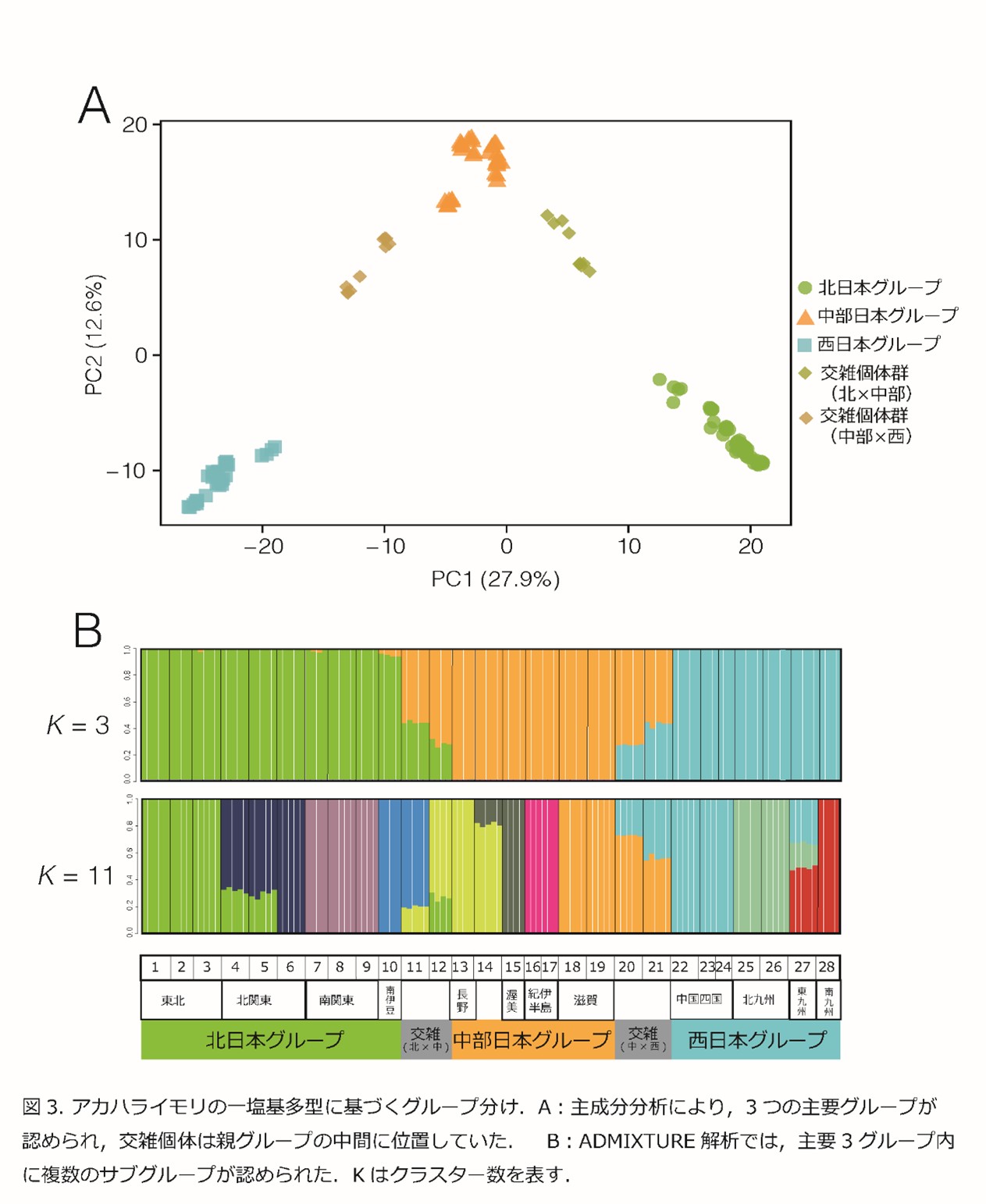
次に,3つの大きなグループの分岐順序を調べるため,ABC解析(*8)を行いました.その結果,まず約820万年前ごろに,本種の共通祖先が北日本グループと西日本グループに分岐した後,約325万年前ごろに両グループの交雑により中部日本グループが生じたことが示されました(図4).820万年前には.現在の関東や東北に相当する多くの地域は海面下にあり,また寒冷化も進行していたことから本種の分布は温暖な地域に限定されていたと考えられます.北日本グループと西日本グループの分岐は,現在の西南日本に存在した複数の孤立した生息適地に集団が隔離されたことで生じたと考えられます.その後,やや温暖化すると,両グループ分布を拡大する過程で,再び分布域が重なり.その過程で交雑が生じ,中部日本グループが生じたものと考えられます.
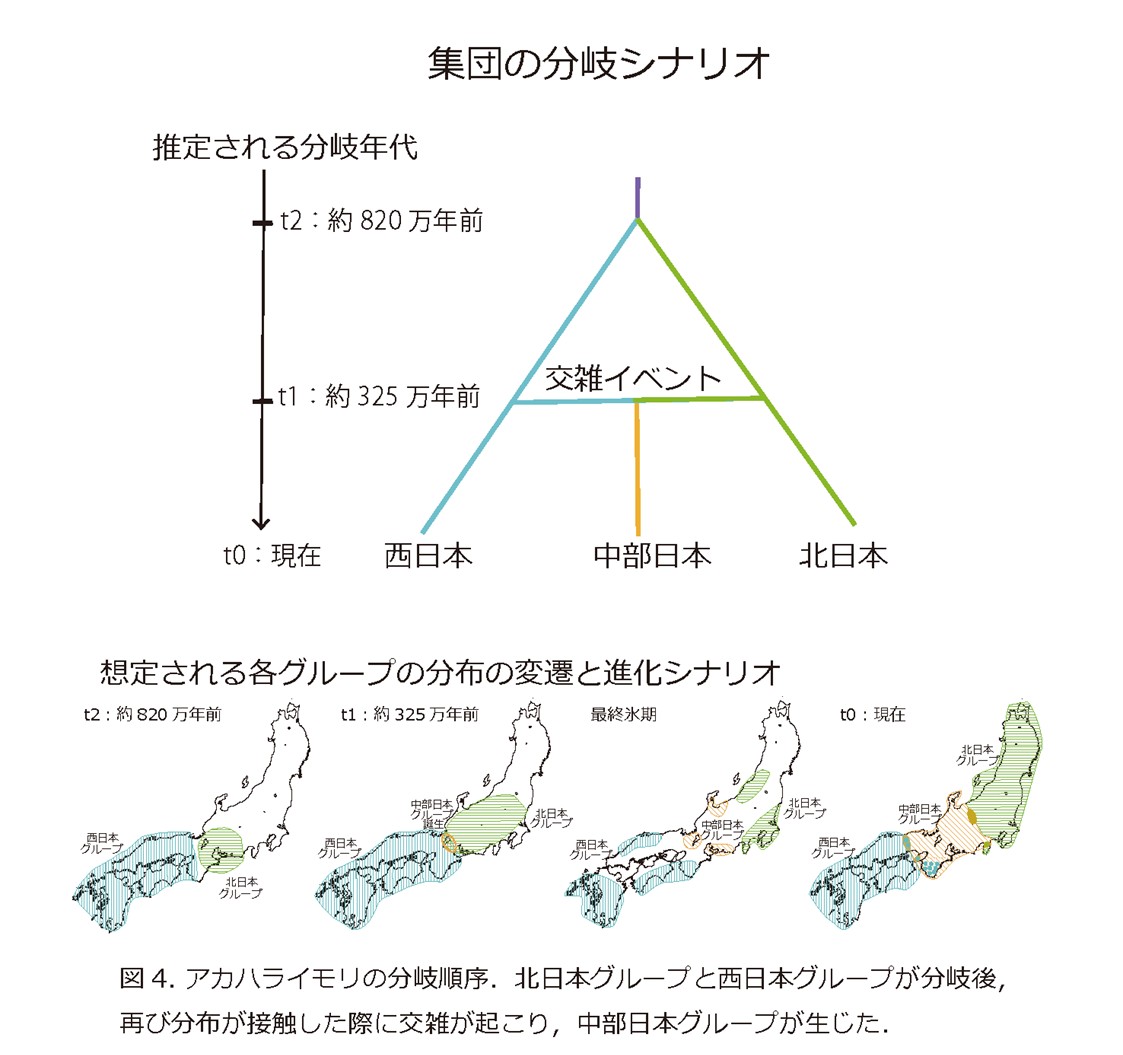
最後に,中部日本グループに対する北日本グループと西日本グループからの過去の遺伝子浸透を調べるため,4集団検定(*9)を行いました.その結果,中部日本グループの西側のサブグループは西日本グループからの遺伝子浸透の影響を受けたのに対し,東側のサブグループは北日本グループからの遺伝子浸透の影響を受けていたことが明らかになりました.また,紀伊半島のサブグループは,ミトコンドリアDNAでは西日本グループに含まれるのに対し,核ゲノムでは中部日本グループに含まれるという,ミトコンドリアDNAと核ゲノムの不一致が確認されました(図2).これらの結果は,過去のグループ間の交雑と分布の変遷が,中部日本グループ内の複雑な集団構造の形成に影響を与えていることを示唆します.
社会的意義・今後の予定
これらの結果は,交雑とその後の遺伝子浸透が,アカハライモリの複雑な集団構造の形成と遺伝的多様性の成立に重要な役割を果たしていることを示しています.アカハライモリのグループ間の遺伝的分化の程度は,別種として分けられてもおかしくない程度の違いにも相当することから,今後はグループ間の分類学的な関係について再検討が必要です.さらに中部日本グループが他の2グループの雑種起源であることを示した本研究の結果は,これまで限られた例しか報告されていない雑種種分化*10)がアカハライモリにおいても生じたことを示唆しており,生物の多様化プロセスの理解を深めるための一助となります.
今後は,交雑によって遺伝的グループを超えて適応的な遺伝子が取り込まれ,グループの分布拡大の足がかりになっている可能性があるのかを調べるため,どのような遺伝子が浸透し,どのような働きを持つのか明らかにしていく必要があります.
<用語解説>
(*1)遺伝的集団構造:同じ種の中で,遺伝的な特徴が地域ごとにどのように分かれて分布しているかを示すもの.地理的な隔離や移動の制限などによって,集団間に遺伝的な違いが生じることで形成される.
(*2) 遺伝子浸透:異なる種や系統間で生じた交雑個体が雑種同士ではなく親系統と戻し交雑を繰り返すことで,一方の系統由来の遺伝子が他方の系統集団へ取り込まれる現象. 種内・系統内の対立遺伝子頻度を変化させることで,その進化に影響を与える要因となる.
(*3)対立遺伝子:同じ遺伝子座(染色体上の同じ位置)に存在する,異なるタイプの遺伝子.
(*4)交雑帯:異なる集団や系統の分布境界で,両者が交配して生じた雑種個体が多く見られる地域.
(*5) ミトコンドリアシトクロムb領域:細胞内の小器官であるミトコンドリアが持つDNAの一部で生物の進化や集団間の遺伝的な違いを調べる研究でよく利用される領域.
(*6) ddRAD-seq:2種類の制限酵素を用いてゲノムを切断し,特定の断片のみを次世代シーケンサーで解析することで多数の一塩基多型 (SNP) 情報を取得する解析手法.
(*7)SNP:遺伝子上に存在する一塩基の違い(多型)のこと.一塩基多型とも呼ばれる.
(*8) ABC解析:近似ベイジアン計算(Approximate Bayesian Computation)を用いてSNPsデータから集団の分岐や交雑の歴史を推定する解析手法.
(*9) 4集団検定:4つの集団の遺伝情報を比較し,異なる集団ペア間の遺伝子の混ざり具合の相対的な大きさを比較することで交雑や遺伝子のやり取りがあったかを調べる解析手法.
(*10)雑種種分化:異なる種同士の交配によって生じた雑種が,独自の特徴や生態を獲得し,親種とは異なる新たな種へと進化する現象.
<論文情報>
(1) Hybridization Drives High Genetic Diversity in the Japanese Fire-Bellied Newt, Cynops pyrrhogaster
(交雑はアカハライモリの高い遺伝的多様性をもたらす)
(2) Journal of Heredity
(3) Hiroki Shiroma*, Masatoshi Matsunami, Tomohiko Shimada, Masafumi Matsui, Kanto Nishikawa, Atsushi J. Nagano, Atsushi Tominaga*
(4) DOI:10.1093/jhered/esag043
(5) URL: https://doi.org/10.1093/jhered/esag043
