|
琉球大学戦略的研究プロジェクトセンターの鶴井香織特命助教および琉球大学農学部の辻瑞樹教授らの琉球大研究チームによる研究成果が生態学・進化学の学術雑誌「Ecology and Evolution」誌に掲載されました。 
【発表のポイント】
|
<発表概要>
【研究背景】
敵の目を欺いて身を守る擬態 「擬態」(ぎたい)とは、天敵に食べられないために、天敵にとって餌でないものに姿や形を似せることです。たとえば天敵が嫌うハチに似た黒と黄色の縞模様の体をしている蛾や、枯れ枝にそっくりなシャクトリムシも擬態していると考えられています。このように、擬態は自然界にごくありふれた現象です。擬態のうち、自分自身は毒を持たない生物が有毒な生物に似せることで天敵から身を守るものを「ベイツ型擬態」と呼びます。ベイツ型擬態の実例は昆虫やヘビなどに見られ、とりわけ蝶でよく知られています。本研究グループも、琉球列島に広く生息するシロオビアゲハというアゲハチョウの一種に注目し、擬態の謎に迫りました。
擬態しない個体の謎 ベイツ型擬態には、「進化論」で有名なダーウィン以来の謎がありました。それは、「擬態する種の中に擬態しない個体が混じっていることがある」ということです。擬態すると天敵に襲われにくくなり得をすると考えられるので、なぜあえて擬態しないのか、とても不思議なことです。研究が進むにつれて、メスは擬態する、オスは擬態しない、と性によって擬態する/しないが決まっている蝶については理由が分かってきました。オスとメスで擬態することの重要さが異なり、メスの方がより擬態する必要性が高いため、メスだけに擬態が進化したのだというわけです。メスは卵を抱えているので餌として栄養豊富で、オスよりも天敵に襲われやすいのです(大崎2009)。
しかし、擬態しない個体の謎はまだ残ります。メスの一部だけが擬態し、その他のメスと全てのオスは擬態しない、という蝶が世界に何種もいるのです。なぜ一部のメスしか擬態しないのか?どれだけのメスが擬態するかはどのように決まるのか?という謎は、長年昆虫学者を魅了してきました。謎を解くカギは、擬態によって蝶がどのように身を守るのか、仕組みの原点に立ち返って考えることにありました。
鳥の学習、毒蝶の数、擬態蝶の数 ベイツ型擬態は、本当は「美味しい」擬態蝶が毒蝶にみせかけて鳥をあざむく現象です。この仕組みは、鳥が毒蝶を食べて、毒蝶の見かけを学習することで成立します。しかし、鳥は、毒蝶にしばらく出会わないと、毒蝶の見かけや不味さを忘れてしまいます。擬態蝶が鳥をあざむく状況は、鳥が頻繁に毒蝶を見たり食べたりして、毒蝶の見た目を忘れないときに起こります。この仕組みを数学を使って考えた研究によれば、擬態蝶が毒蝶に比べて増え過ぎると、鳥の擬態学習効率が悪くなり、擬態しても食べられやすくなることが予測されました(Barrett 1976)。この予測はつまり、「毒蝶の数が多いほど擬態できる蝶の数も多くなる」というものです。この予測を「擬態率の毒蝶依存説」と呼ぶことにします。擬態率は、集団中の何割が擬態するかを意味します。
【研究内容】
研究のアイデア:琉球列島のシロオビアゲハで仮説証明 「擬態率の毒蝶依存説」を証明するためには毒蝶の数が異なる地域間で毒蝶に対する擬態蝶の割合を比較する必要があります。琉球大学戦略的研究プロジェクトセンターの鶴井香織特命助教、佐藤行人特命講師、および琉球大学農学部の辻瑞樹教授らの研究チームは、琉球列島の島々に広く生息している黒いアゲハチョウ、シロオビアゲハに注目しました(図1a)。シロオビアゲハはメスの一部だけがベニモンアゲハという毒蝶(図1a)に擬態する蝶です。研究グループは、琉球列島に散らばるベニモンアゲハの多い島と少ない島(図1b)の間でシロオビアゲハの擬態率を比較して「擬態率の毒蝶依存説」を実証することにしました。つまり、ベニモンアゲハが多い島ほど、鳥が毒蝶の見た目を「良く覚えている」ので、多くのシロオビアゲハのメスが擬態によって身を守ることができ、シロオビアゲハのメスの擬態率が結果的に高くなっているだろうという考えです。実は、1982年に上杉兼司氏が琉球列島の島々でシロオビアゲハとベニモンアゲハの個体数を観測したデータ(上杉2000)は、ベニモンアゲハが多い島ほどシロオビアゲハの擬態率が高くなる様子を見事にとらえていました。しかし、これ以来40年近くもの間、野外での再検証はなされませんでした。そこで研究グループは、上杉氏による1982年の調査結果を再検証し、さらに、その結果が鳥による捕食以外の要因(島ごとの蝶の遺伝的な関係など)に左右されていないことをDNA解析により裏付けることにしました。
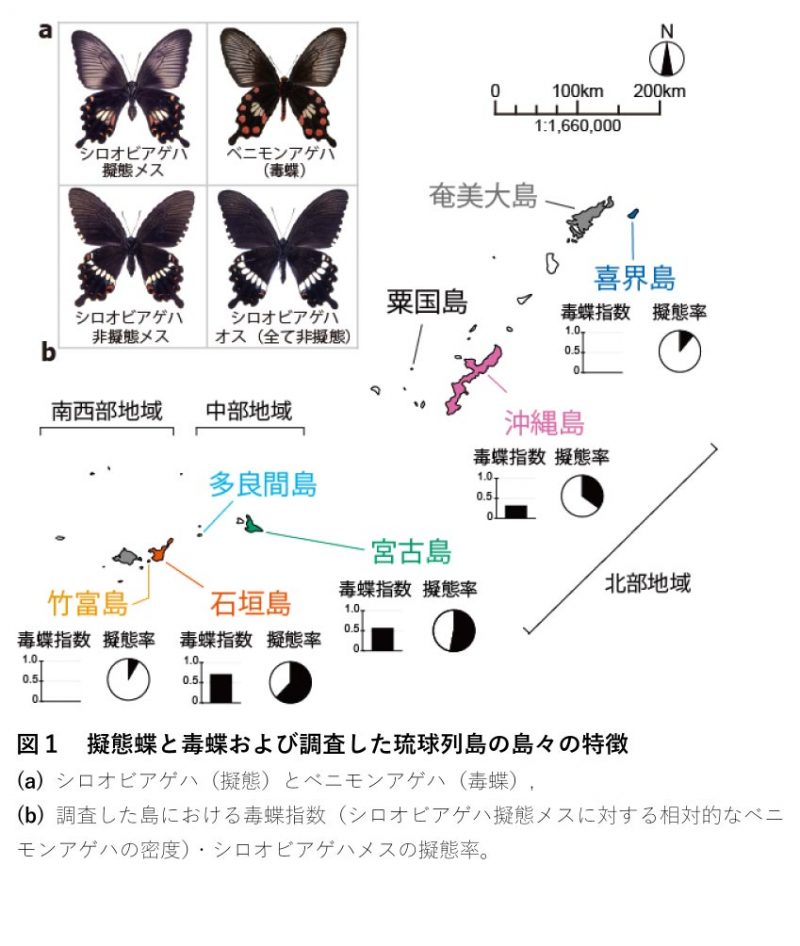
野外調査 研究グループは、2014~2017年に、琉球列島の喜界島・沖縄島・宮古島・石垣島・竹富島の5つの島で、ベニモンアゲハとシロオビアゲハの個体数と擬態を確認する野外調査を行ないました。そして、毒蝶であるベニモンアゲハが多い島ほどシロオビアゲハの擬態率が高くなるか(図2において、毒蝶指数が高いほどシロオビアゲハの擬態率が高くなるか)を統計的に解析しました(注1)。結果は我々の予測通り、シロオビアゲハの擬態率は毒のあるベニモンアゲハが多い島ほど高くなりました(図2)。そしてこの結果は、上杉氏による1982年の調査結果を再現するものでした。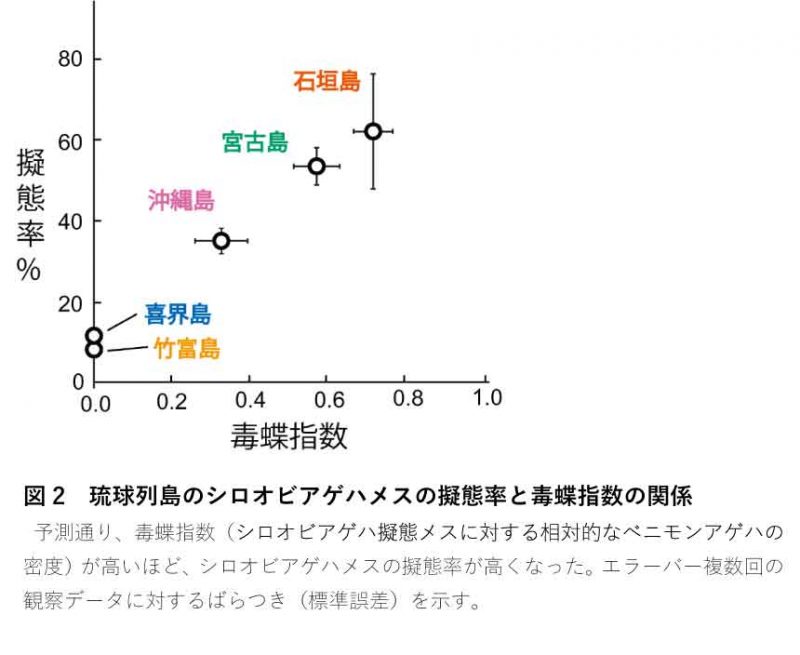
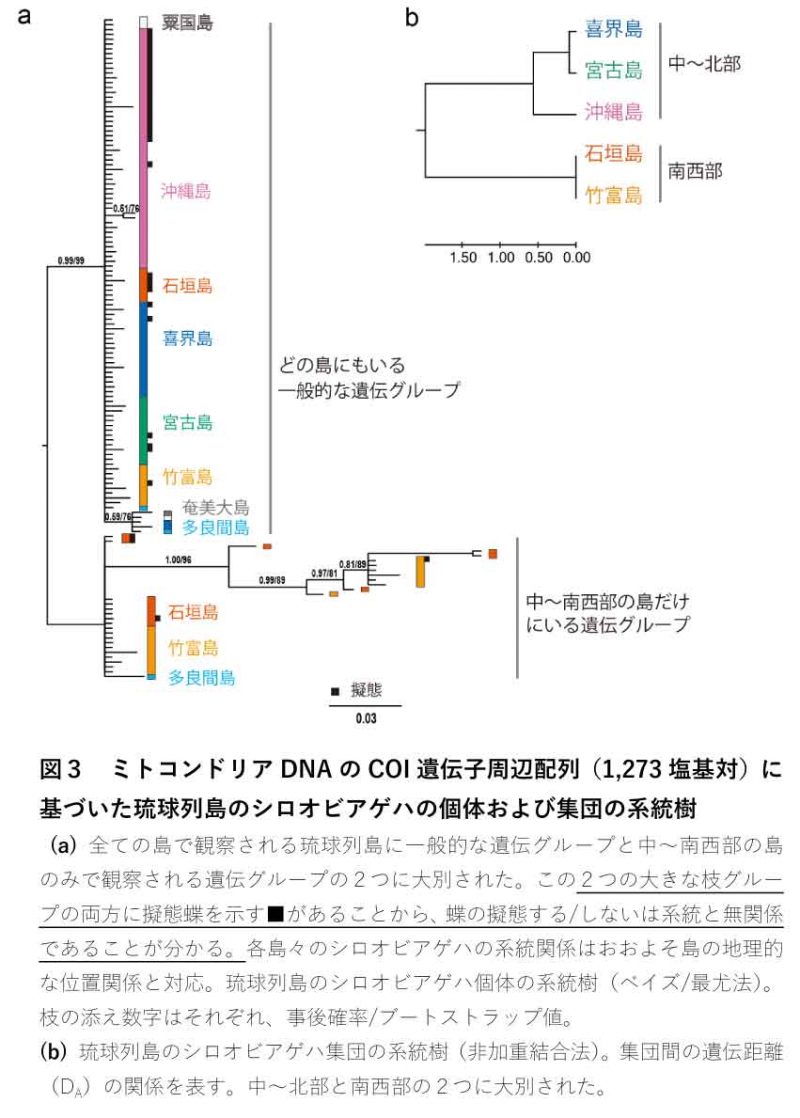
ミトコンドリアDNA解析 「擬態率の毒蝶依存説」はいわゆる自然選択説に基づいた説明です。しかし、一方で、シロオビアゲハの擬態率を決める要因として、自然選択以外の可能性も考えられます。シロオビアゲハの擬態率は毒蝶とは関係なく、たまたま島に定着した集団の家系がもともと持っていた遺伝的な擬態しやすさの影響を受ける可能性があります。また、擬態率の高い隣接した島からシロオビアゲハが頻繁に移住してくると、それによって擬態率が上がる可能性もあります。そこで、このような偶然の出来事がシロオビアゲハの擬態率に影響する可能性について、次世代シーケンサー(注2)で解読したミトコンドリアDNA(注3; 以降mtDNAと表記)配列を解析することで検証しました。調べたのは、琉球列島の喜界島・沖縄島・宮古島・石垣島・竹富島を中心に、奄美大島・粟国島・多良間島も含めた8つの島で採集したシロオビアゲハのDNAです。
系統関係と擬態率 シロオビアゲハmtDNAにあるCOI遺伝子周辺の塩基配列(1,273塩基対)に基づいた137匹のシロオビアゲハの系統解析の結果、8つの島で採集されたシロオビアゲハの系統関係はおおよそ、大きな2つの遺伝的なグループに分かれましたが(図3a, b)、擬態する/しないは系統とは無関係であることが分かりました(図3aの■)。さらに、70匹のシロオビアゲハについて同じくmtDNAにあるCOIII遺伝子とCytb遺伝子周辺領域の塩基配列(それぞれ442・568塩基対)のデータを追加して系統解析を行ないましたが、結果は同じでした。
集団の歴史と擬態率 mtDNAの配列情報をもとに、島に定着したシロオビアゲハの集団が存続している時間をおおよそ推定できます。この推定によると、石垣島と竹富島のシロオビアゲハ集団はかなり古くからこれらの島に定着していたと考えられます。一方で、喜界島と宮古島の集団は比較的新しい集団であると推定されました。シロオビアゲハの擬態率は石垣島や宮古島で高く、喜界島や竹富島で低い(図1, 2)ため、シロオビアゲハの集団の古さも擬態率とは関係ないことが分かります。
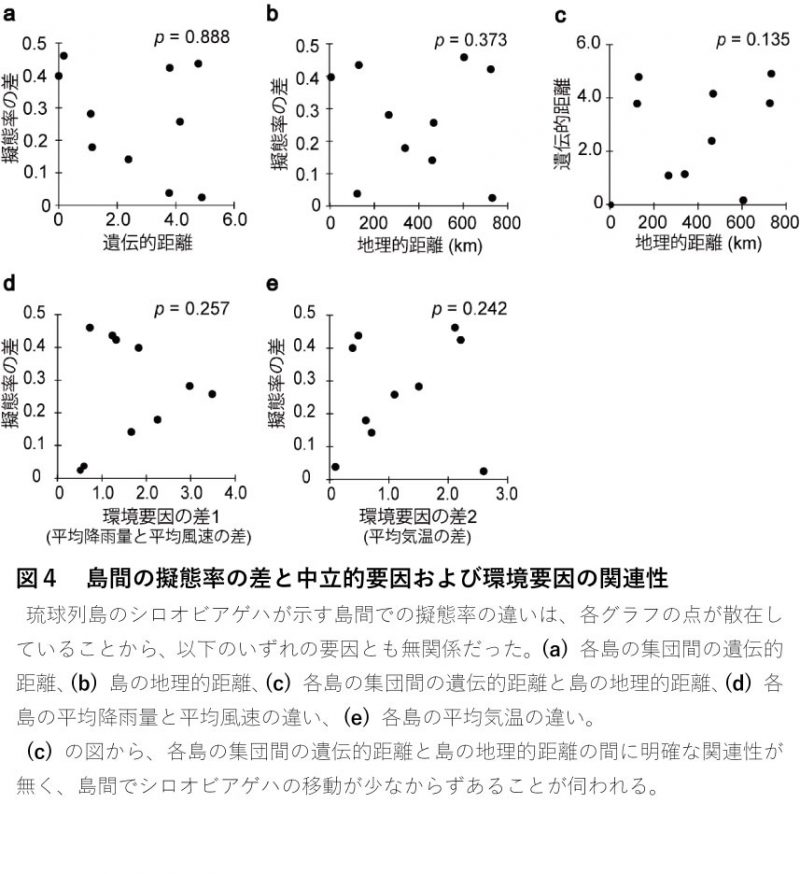
遺伝的距離・地理的距離・気候環境の違い 最後に、シロオビアゲハmtDNAの配列情報や気象庁のアメダスデータをもとに、島間の擬態率の差が遺伝的距離、地理的距離、気候環境の違いと関連しているか調べましたが、どの要因も擬態率の違いに寄与していませんでした(図4)。つまりシロオビアゲハの擬態率は、個体の移動などの要因や気候環境の違いにほとんど左右されないことが分かりました。
結論 以上を総合し、シロオビアゲハの擬態率が毒蝶の多い島ほど高いのは、天敵である鳥の学習を介した自然選択の作用によるものであり、蝶の遺伝系統や移動分散、島の気候環境とは関係ないと結論付けました。つまり、シロオビアゲハの擬態率がモデルとなる毒チョウの比率に依存して変化するという仮説が支持されました。これは、「擬態率の毒蝶依存説」が支持された世界初の例です。
【社会的意義・今後の予定等】
40年間での擬態率の急速変化 研究チームにより、「毒蝶が多い地域ほど擬態できる蝶の数も多くなる」という「擬態率の毒蝶依存説」が実証されました。もう1点、研究チームが過去のデータを再検証したことで、新たに面白い傾向が見つかりました。研究チームのデータと1982年のデータを比較すると、約40年間の間に、石垣島・宮古島では毒蝶のベニモンアゲハの数が増え、それに伴い、シロオビアゲハの擬態率もそれぞれ30%程度上昇していることが分かりました。つまり、40年という短い間に、シロオビアゲハの擬態率は、毒蝶の相対密度に応じた鳥の学習と捕食によって島ごとに個別に急速進化したということです。
琉球列島で起こるリアルタイムな擬態の進化 実は、毒蝶のベニモンアゲハは、もともとは台湾以南に生息する熱帯の種で比較的最近まで琉球列島に生息してはいませんでした。ところが、石垣島には1968年、宮古島には1975年、沖縄本島には1993年に移動・定着したと報告されています。この数十年で琉球列島の各島のベニモンアゲハが急激に増え、シロオビアゲハをとりまく擬態の状況も刻々と変化しています。今、琉球列島のシロオビアゲハは、リアルタイムで擬態が急速に進化していると言えます。2017年には、本研究チームのメンバーである大学院生の加藤三歩さんと辻教授らがおこなった研究(Katoh et al. 2017)で、シロオビアゲハの翅の擬態模様のうち後翅の白い斑紋(白紋)が、この数十年で有毒なベニモンアゲハに似るように急速に進化したことが明らかになっています。琉球列島のシロオビアゲハは擬態の進化をリアルタイムで観察できる世界でも極めて珍しく貴重な存在なのです。
擬態の進化研究で世界から注目のシロオビアゲハ シロオビアゲハは、沖縄では最も身近でありふれたアゲハチョウですが、世界中の擬態研究者、とりわけ擬態遺伝子研究者から最も注目を集めているチョウの1つで、野生の昆虫としては遺伝学の知見が多く蓄積されています。沖縄ではシロオビアゲハが身近なため、シロオビアゲハの擬態に関連する行動や生態を直接観察する機会に恵まれています。私たち研究チームはこうした地の利を活かし、行動・生態・遺伝といった様々な視点から、更なる研究を実施していく予定です。
<用語解説>
(注1)ベニモンアゲハの多さは、シロオビアゲハ擬態メスとの相対的な割合を示す「毒蝶指数」として以下の計算式に基づいて評価しました。
また、シロオビアゲハの擬態率は、以下の式で計算しました。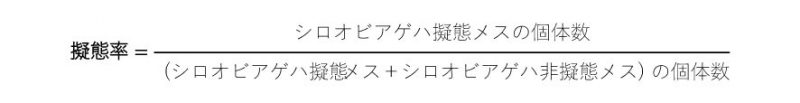
(注2)遺伝子の塩基配列を高速に読み出せる装置
(注3)細胞小器官であるミトコンドリア内にあるDNAのことです。ミトコンドリアDNAの塩基配列を解読するための技術開発が進んでいるため、塩基配列情報に乏しい野生生物においても、ミトコンドリアDNAの塩基配列の解析は比較的実施しやすいとされています。そのため、野生生物を対象とした生態学的研究における遺伝解析ではミトコンドリアDNAがよく利用されます。
<引用文献>
Barrett, J. A. (1976). The maintenance of non‐mimetic forms in a dimorphic Batesian mimic species. Evolution, 30(1), 82–85.
Katoh M et al. (2017) Rapid evolution of a Batesian mimicry trait in a butterfly responding to arrival of a new model. Scientific Reports 7, Article number: 6369.
大崎直太 (2009)『擬態の進化. ダーウィンも誤解した150年の謎を解く』, 海游舎.
上杉兼司(2000)成虫はどうやって身を守っているのか. 擬態と多型. 『蝶の自然史-行動と生態の進化学-』(大崎直太 編著), 北海道大学図書刊行会, pp106-123.
<論文情報>
(1)タイトル
Evidence for frequency‐dependent selection maintaining polymorphism in the Batesian mimic Papilio polytes in multiple islands in the Ryukyus, Japan
(和訳)負の頻度依存選択がベイツ型擬態多型を維持している証拠:琉球列島のシロオビアゲハを用いた実証
(2)雑誌名
Ecology and Evolution
(3)著者
鶴井香織*,†,1 佐藤行人†,1 加藤絵美†, 2 加藤三歩†, 2, 3 木村亮介4 立田晴記2, 3 辻和希(瑞樹)*2, 3
* Corresponding author
†These authors contributed equally to this work.
1 琉球大学戦略的研究プロジェクトセンター
2 琉球大学農学部/琉球大学大学院農学研究科
3 鹿児島大学連合大学院連合農学研究科
4 琉球大学医学研究科
(4)DOI 10.1002/ece3.5182
(5)論文(アブストラクトを含む全文)URL
https://onlinelibrary.wiley.com/doi/full/10.1002/ece3.5182
